
By Jessica Brösamlen
Daily actions and thoughts emerge as cells in the central nervous system are constantly sending molecular messengers to adjacent neurons and in this manner executing a steady informational transfer. Glial cells, surrounding neuronal cells, play a crucial role in forming and maintaining the Blood Brain Barrier. Their functions range from homeostatic regulators (astrocytes) to trash collectors (microglia). Like all cells in our body, their specific role is encoded in our DNA and is regulated by gene expression. Nonetheless, small DNA changes (mutations) by cause of internal (inherited) or external (environmental) factors may underlie susceptibility to various disorders, one of which is Parkinson’s Disease (PD). PD is the second most common neurodegenerative disorder, affecting mainly people above the age of 60. More than two centuries have passed since the first medical case was described1 and surprisingly we still don’t have curative treatments. One plausible explanation is the focus of research for many decades on neurons and the lack of understanding of the importance of glia cells (astrocytes and microglia).
The molecular mechanism of PD
There are two significant hallmarks of PD. Firstly, the loss of neurons in an anatomical pathway which takes part in dopamine production is called the nigrostriatal pathway. Dopamine signalling here is crucial for initiating and modulating movement in our bodies.
These neurons contain a black pigment protein called Neuromelanin (NM), which can help in the removal of excessive amounts of dopamine or other toxins in the cell. Interesting to note is that this protein is released when the neurons die. Secondly, PD patients present intraneuronal inclusions, known as Lewy body aggregations. These are mainly composed of the protein alpha-synuclein (α-syn) (Figure B)5. Alpha-synuclein found outside the cells as well as NM aggregates are considered to cause neurotoxicity via an inflammatory pathway 6,7.

Shifting the focus to glia cells might help unravel the mechanism behind PD
Traditional research has mainly been focused on the degenerating dopamine neurons, disregarding other cell types in the central nervous system (CNS) that could also contribute to PD. The interest in unravelling this possible disruption of the multi-cellular interaction between different CNS-specific cell types appeared only after researchers found reactive support cells, known as glia cells (microglia and astrocytes) in post-mortem brain samples.
Lack of early diagnostic tools and curative treatment in PD

Prominent clinical symptoms in PD are motor problems, such as rigidity, postural instability and tremors at rest2 and non-motor symptoms, such as sleep loss, olfactory loss and cognitive disruptions 3. In the majority of cases, the latter occurs before the onset of motor symptoms. and patients are rarely diagnosed before the motor symptoms present. By this stage, the neuronal loss is typically too far advanced.
Currently, pharmaceutical medications, such as Levodopa, work to restore dopamine levels after neuron loss, and therefore represent only a symptomatic and not a curative treatment of PD. Disease-modifying pharmacological treatments that stop neuronal degeneration and early diagnostic tools are crucially missing. In this way, there might be further hope for a novel treatment, primarily controlling the toxic activity of cell types surrounding the dying neurons.
Mutations in immune-related genes and PD
There are various mutated genes linked to PD, one of which is the leucine-rich-repeat kinase 2 (LRRK2),. This gene has been described to be involved in immunity. The study that shed light on the close link between LRRK2 and microglia illustrated that the over-expression of the LRRK2 protein caused microglia activation which subsequently triggered neuro-inflammation6.

Microglia and Astrocytes show bipolar phenotypes: How can they be Neuro-supportive and Neurotoxic?
Microglia (MG) are the immune cells of the CNS. They work by stretching out and retracting their processes to constantly surveille the local environment. They act upon inflammation by releasing messengers, known as cytokines which lead to a wide range of actions from synapse rewiring during neuronal development to neuronal death when toxins are present8.
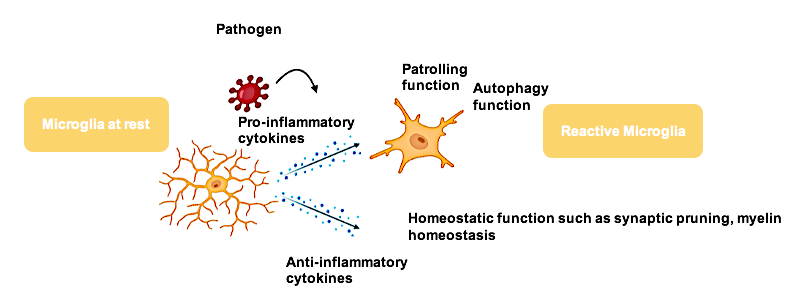
On the other hand, astrocytes nourish neurons and are responsible for maintaining a homeostatic environment. They are the most abundant cells in the brain that are in direct contact with the blood-brain barrier and surrounding neurons, taking up not only neuronal released ions but also toxins that further cause astrocytic pro-inflammatory cytokine release.
Both types of glia cells, microglia and astrocytes are essential to maintain balance in the system. When exposed to inflammatory stimuli that trigger instability, they rapidly modulate their resting state towards an active state. Microglia and astrocytes operate in a non-controlled manner when they carry mutations in genes that are vital for immune regulation. This chronic activity may lead to the harmful switch from being protective cells to becoming disruptive, finally causing neuronal degeneration7,9,10,11.
The link between LRRK2 – Glia cell – Inflammation – Neurodegeneration in PD
The CNS is constantly being exposed to stimuli (i.e. pathogen (external) or protein aggregates (internal) that may be malignant and cause activation of the immune system. The first responders are the immune cells, microglia, that release cytokines upon activation. These in turn cause the activation of astrocytes which increases the release of pro-inflammatory cytokines, subsequently leading to the death of toxic neurons.
In PD, the release of NM or alpha-synuclein aggregates triggers the activation of glial cells to phagocytose toxins and protect surrounding neurons. However, if microglia- and astrocytic functions are disrupted by a mutation such as LRRK2, the sustained inflammatory pathway will cause a spiral reaction inducing unstoppable neuronal death. Therefore, it is imperative to further explore this neuron-glial interaction further to continue the search for a cure for Parkinson’s.
About the author

Jessica Brösamlen is a second-year neuroscience master’s student, studying the effects of Glia cells in Parkinson’s Disease using induced pluripotent stem cell technology. Specifically, she works on introducing all three cell types (microglia, astrocytes and neurons) in a culture dish and studying their interaction.
Further Reading
- Pons-Espinal, M., Blasco-Agell, L., & Consiglio, A. (2020). Dissecting the non-neuronal cell contribution to Parkinson’s disease pathogenesis using induced pluripotent stem cells. Cellular And Molecular Life Sciences, 78(5), 2081-2094. doi: 10.1007/s00018-020-03700-x
- Olanow, C. W., Stern, M. B., & Sethi, K. (2009). The scientific and clinical basis for the treatment of Parkinson disease (2009). Neurology, 72(21 Suppl 4), S1–S136. https://doi.org/10.1212/WNL.0b013e3181a1d44c
- Emre, M., Aarsland, D., Brown, R., Burn, D. J., Duyckaerts, C., Mizuno, Y., Broe, G. A., Cummings, J., Dickson, D. W., Gauthier, S., Goldman, J., Goetz, C., Korczyn, A., Lees, A., Levy, R., Litvan, I., McKeith, I., Olanow, W., Poewe, W., Quinn, N., … Dubois, B. (2007). Clinical diagnostic criteria for dementia associated with Parkinson’s disease. Movement disorders : official journal of the Movement Disorder Society, 22(12), 1689–1837. https://doi.org/10.1002/mds.21507
- Postuma, R. B., Aarsland, D., Barone, P., Burn, D. J., Hawkes, C. H., Oertel, W., & Ziemssen, T. (2012). Identifying prodromal Parkinson’s disease: pre-motor disorders in Parkinson’s disease. Movement disorders : official journal of the Movement Disorder Society, 27(5), 617–626. https://doi.org/10.1002/mds.24996
- Braak, H., & Braak, E. (1995). Staging of Alzheimer’s disease-related neurofibrillary changes. Neurobiology of aging, 16(3), 271–284. https://doi.org/10.1016/0197-4580(95)00021-6
- Hirsch, E., Hunot, S., Damier, P., & Faucheux, B. (1998). Glial cells and inflammation in parkinson’s disease: A role in neurodegeneration?. Annals Of Neurology, 44(S1), S115-S120. doi: 10.1002/ana.410440717
- Abbott A. (2010). Levodopa: the story so far. Nature, 466(7310), S6–S7. https://doi.org/10.1038/466S6a
- Zucca, F. A., Segura-Aguilar, J., Ferrari, E., Muñoz, P., Paris, I., Sulzer, D., Sarna, T., Casella, L., & Zecca, L. (2017). Interactions of iron, dopamine and neuromelanin pathways in brain aging and Parkinson’s disease. Progress in neurobiology, 155, 96–119. https://doi.org/10.1016/j.pneurobio.2015.09.012
- Wake, H., Moorhouse, A. J., Jinno, S., Kohsaka, S., & Nabekura, J. (2009). Resting microglia directly monitor the functional state of synapses in vivo and determine the fate of ischemic terminals. The Journal of neuroscience : the official journal of the Society for Neuroscience, 29(13), 3974–3980. https://doi.org/10.1523/JNEUROSCI.4363-08.2009
- Langston, J. W., Forno, L. S., Tetrud, J., Reeves, A. G., Kaplan, J. A., & Karluk, D. (1999). Evidence of active nerve cell degeneration in the substantia nigra of humans years after 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine exposure. Annals of neurology, 46(4), 598–605. https://doi.org/10.1002/1531-8249(199910)46:4<598::aid-ana7>3.0.co;2-f
- Zhang, W., Wang, T., Pei, Z., Miller, D. S., Wu, X., Block, M. L., Wilson, B., Zhang, W., Zhou, Y., Hong, J. S., & Zhang, J. (2005). Aggregated alpha-synuclein activates microglia: a process leading to disease progression in Parkinson’s disease. FASEB journal : official publication of the Federation of American Societies for Experimental Biology, 19(6), 533–542. https://doi.org/10.1096/fj.04-2751com
- Hickman, S., Izzy, S., Sen, P., Morsett, L., & El Khoury, J. (2018). Microglia in neurodegeneration. Nature Neuroscience, 21(10), 1359-1369. doi: 10.1038/s41593-018-0242-x
- di Domenico, A., Carola, G., Calatayud, C., Pons-Espinal, M., Muñoz, J. P., Richaud-Patin, Y., Fernandez-Carasa, I., Gut, M., Faella, A., Parameswaran, J., Soriano, J., Ferrer, I., Tolosa, E., Zorzano, A., Cuervo, A. M., Raya, A., & Consiglio, A. (2019). Patient-Specific iPSC-Derived Astrocytes Contribute to Non-Cell-Autonomous Neurodegeneration in Parkinson’s Disease. Stem cell reports, 12(2), 213–229. https://doi.org/10.1016/j.stemcr.2018.12.011
Image Credits: Cover photo by Jessica Brösamlen; Figures 1A, 2, 3, 4, 5 were created using Biorender; Figure 1B is by Tulemo – Own work, CC BY-SA 4.0; Figure 2 is adapted from Arora et al 2022